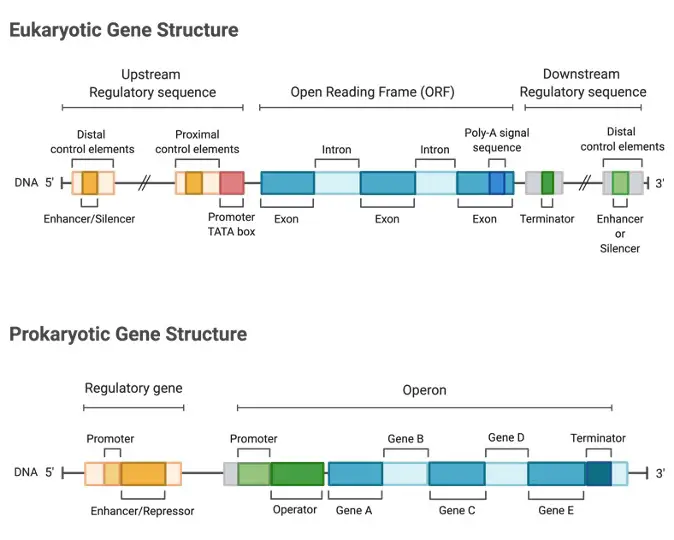
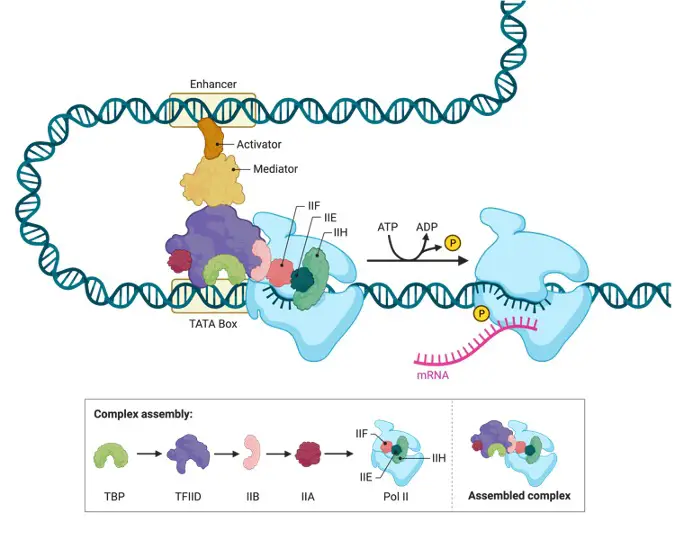
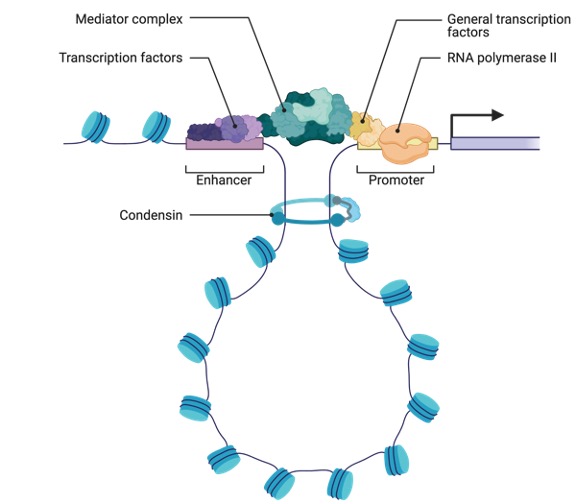
MyoD is a transcription factor that can cause muscle tissue to differentiate. It’s just one of many regulators involved in muscle development, but it’s plenty to show how complicated eukaryotic gene control can be. Skeletal muscle is a multinucleate tissue made up of embryonic myoblasts that have fused together.
MyoD, Myf5, Myogenin, and MRF4 are four key regulators of muscle cell development. Mice lacking either MyoD or Myf5 create normal muscle tissue, whereas mice lacking both MyoD and Myf5 fail to generate any skeletal muscle at all, indicating that these two genes regulate muscle development jointly.
Mice lacking the other two genes, on the other hand, have a variety of muscular impairments, indicating that all four genes are necessary. MyoD binds to the regulatory areas of several of the genes involved in skeletal muscle formation.
It generally does not operate alone, but rather forms heterodimers with other proteins that have comparable functions. These dimers detect pairs of E-box DNA regulatory elements, the consensus sequence of which is CANNTG.
MyoD may also play a role in muscle cell differentiation by attaching to an enzyme that acetylates histones, which allows it to rearrange the chromosomes around muscle-specific genes, making them accessible to transcription. Histones are attracted to the negatively charged phosphate groups of the DNA backbone because they are basic proteins.
Acetylating histones lowers their affinity for DNA, allowing it to be released for transcription. The functions of MyoD in activating muscle-specific genes are oversimplified, and MyoD itself has many regulators. It is, however, a brief glimpse into the intricacies of eukaryotic transcriptional control.